Muitos trabalhos foram escritos sobre a importância da glicose 6 fosfato desidrogenase na proliferação e na morte celular. Na via das pentoses, ela é a enzima limitante do ramo oxidativo e sua principal função é gerar NADPH. No ramo não oxidativo a enzima limitante é a transcetolase cuja principal função é produzir ribose, coluna dorsal do DNA e RNA.
Na via das pentoses a ativação da glicose 6 fosfato desidrogenase produz uma molécula de NADPH e a ativação da enzima subseqüente a fosfoglucodehidrogenase produz outra molécula de NADPH e assim cada mol de glicose produz dois moles de NADPH.
O NADPH produzido na via das pentoses é o principal agente redutor intracelular. Ele é o principal fornecedor de átomos de hidrogênio (elétrons) no citoplasma e o seu papel fundamental é manter a glutationa em seu estado reduzido (GSH) o que protege os grupos sulfidrilas e a integridade celular do excesso de radicais livres de oxigênio.
De fato, quando o meio intracelular é redutor, isto é, o equilíbrio da oxi-redução tende para a redução (excesso de antioxidantes, excesso de agentes doadores de hidrogênio ou de elétrons), à medida que a GSSG (glutationa oxidada) vai sendo formada ela é reduzida para GSH a qual ativa a glicolíse anaeróbia que é o motor da mitose, aumentando a proliferação celular neoplásica. Este meio redutor facilita a fosforilação da proteina retinoblastoma, do NF-kappaB e do importante fator de proliferação celular MAPK, contribuindo para a proliferação celular.
Quando o meio intracelular é oxidante, isto é, o equilíbrio da oxi-redução tende para a oxidação (excesso de oxidantes, excesso de agentes aceptores de hidrogênio ou de elétrons), à medida que a GSSG (glutationa oxidada) é formada ela inibe a glicólise anaeróbia. A inibição da glicólise anaeróbia faz parar o ciclo celular e a conseqüência é a diminuição da proliferação celular neoplásica, com apoptose da célula tumoral. (Felippe-2004).Se o meio intracelular é mantido oxidante, consegue-se bloquear a proliferação celular maligna e a célula pode entrar na fase G0 ou sofrer citotoxicidade, posteriormente caminhando para apoptose e /ou necrose.É muito interessante saber que as células cancerosas requerem apenas um leve aumento do potencial redox para cessarem a proliferação, entretanto este leve aumento deve ser contínuo e ininterrupto até a ocorrência da apoptose, pois se houver queda do potencial redox restaura-se a fosforilação da proteína retinoblastoma e as células voltam a proliferar (Felippe -2004-2005).
O principal agente redutor do intracelular é o NADPH
Antigamente, pensava-se que a glutationa reduzida (GSH) era o principal agente redutor citoplasmático. Atualmente sabemos que o principal agente redutor do citoplasma da maioria das células é o NADPH. A glicose 6 fosfato desidrogenase é que determina os níveis de NADPH controlando a entrada da glicose-6-fosfato na via das pentoses .
A glicose 6 fosfato desidrogenase desempenha papel crítico na proliferação celular, via regulação do potencial redox (Tian-1998). O aumento da atividade da glicose 6 fosfato desdrogenase estimula a proliferação celular verificado pelo aumento da incorporação de timidina H3, enquanto que a sua inibição abole a incorporação de timidina tritiada (Tian-1998). É a falta de NADPH e não a falta de ribose-5-fosfato a responsável pela supressão da proliferação celular causada pela inibição da glicose 6 fosfato desidrogenase.
Ativação da glicose 6 fosfato desidrogenase aumenta a proliferação celular
Muitos modelos diferentes de proliferação celular sugerem que o aumento da atividade da glicose 6 fosfato desidrogenase desempenha papel importante na proliferação celular. Sabe-se que células cancerosas in vivo e células transformadas em cultura apresentam aumento significante da atividade da glicose 6 fosfato desidrogenase em níveis de até 20 vezes maiores que as correspondentes células não cancerosas ou não transformadas (Weber-1987 in Tian-1999). A concentração desta enzima está drasticamente elevada nos tumores metastáticos de fígado em relação às outras enzimas da via das pentoses (Geertrudia -1993).
Resumo dos mecanismos de ação da glicose-6-fosfatodesidrogenase
A - Ativação da enzima:
1-Aumenta a produção de NADPH, diminui o potencial redox intracelular e provoca aumento da proliferação celular maligna, com diminuição da apoptose
2-Eleva a produção de ribose, coluna dorsal do DNA e RNA das células malignas
3-Permite o efeito dos fatores de crescimento tumoral: IGF-I, Insulina, EGF, PDGF
4-Aumenta a resistência à quimioterapia e radioterapia
5-Diminui o efeito dos oxidantes na apoptose e na proliferação celular
6-Ativa a MAPK
7-Ativa a fosforilação da tirosina
B - Inibição da enzima:
1-Diminui a produção de NADPH, aumenta o potencial redox intracelular e provoca inibição da proliferação celular maligna, com aumento da apoptose
2-Diminui a produção de ribose, matéria prima para construção do DNA e RNA
3-Diminui o efeito dos fatores de crescimento tumoral: IGF-I, Insulina, EGF, PDGF
4-Aumenta o efeito da quimioterapia e da radioterapia
5-Aumenta o efeito dos oxidantes na apoptose e na inibição da proliferação celular
6-Inibe a MAPK
7-Inibe a fosforilação da tirosina
domingo, 9 de dezembro de 2007
Ribose e os exercícios físicos
A importância das vias bioenergéticas para contração muscular esquelética tem sido amplamente apreciada nos dias atuais. Contrações musculares intensas e repetitivas geram uma condição de hipóxia no músculo esquelético, ou seja, diminuição na chegada de oxigênio à célula muscular. O corpo então utiliza vias metabólicas anaeróbias na tentativa de sustentar a demanda exigida, através do catabolismo de compostos de fosfatos altamente energéticos como ATP. Porém, como os níveis de oxigênio estão limitados, as contrações musculares podem ser mantidas somente por um curto período de tempo, pois os compostos energéticos disponíveis não são suficientemente capazes de manter as contrações por um tempo prolongado.
Assim como os baixos níveis de ATP, a concentração de adenina nucleotídeos é similarmente afetada durante o exercício intenso, se tornando reduzida durante períodos de hipóxia ou metabolismo anaeróbio. A depleção das moléculas de adenina nucleotídeos e a diminuição de sua concentração podem desencadear o estado de fadiga metabólica e, para recomposição de seus níveis, o corpo utiliza o metabolismo de novo de nucleotídeos purínicos, ou então, as vias de recuperação.
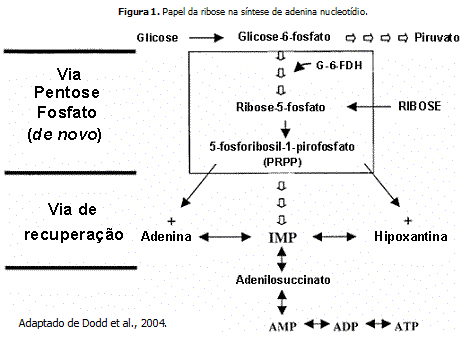
Algumas evidências indicam que um dos principais caminhos para a síntese de novo de nucleotídeos é a fosforilação da ribose em ribose-5-fosfato a partir da via da pentose fosfato. Outros dados indicam que a glicose, através da mesma via, possa ser uma peça chave para o processo regenerativo das moléculas de adenina nucleotídeos, pois a glicose-6-fosfato também pode ser convertida em ribose-5-fosfato. Após essa conversão, a ribose-5-fosfato sofreria uma ação enzimática e resultaria na formação de um novo composto, o 5-fosforibosil-1-pirofosfato (PRPP). Uma vez formado, o PRPP é convertido indiretamente para inosina monofosfato (IMP) que, então, é convertida para adenilosuccinato pela anedilosuccinato sintetase e, em seguida, em AMP, através da adenilosuccinase. Sendo assim, o AMP estará disponível para ser refosforilado para reposição das concentrações de ATP depletadas durante a contração muscular.
Assim como os baixos níveis de ATP, a concentração de adenina nucleotídeos é similarmente afetada durante o exercício intenso, se tornando reduzida durante períodos de hipóxia ou metabolismo anaeróbio. A depleção das moléculas de adenina nucleotídeos e a diminuição de sua concentração podem desencadear o estado de fadiga metabólica e, para recomposição de seus níveis, o corpo utiliza o metabolismo de novo de nucleotídeos purínicos, ou então, as vias de recuperação.
Algumas evidências indicam que um dos principais caminhos para a síntese de novo de nucleotídeos é a fosforilação da ribose em ribose-5-fosfato a partir da via da pentose fosfato. Outros dados indicam que a glicose, através da mesma via, possa ser uma peça chave para o processo regenerativo das moléculas de adenina nucleotídeos, pois a glicose-6-fosfato também pode ser convertida em ribose-5-fosfato. Após essa conversão, a ribose-5-fosfato sofreria uma ação enzimática e resultaria na formação de um novo composto, o 5-fosforibosil-1-pirofosfato (PRPP). Uma vez formado, o PRPP é convertido indiretamente para inosina monofosfato (IMP) que, então, é convertida para adenilosuccinato pela anedilosuccinato sintetase e, em seguida, em AMP, através da adenilosuccinase. Sendo assim, o AMP estará disponível para ser refosforilado para reposição das concentrações de ATP depletadas durante a contração muscular.
Na via da pentose fosfato existe a participação de enzimas como a glicose-6-fosfato desidrogenase e a 6-fosfogluconato desidrogenase, sendo que ambas possuem uma atividade limitada, tanto no músculo esquelético quanto no músculo cardíaco. Um outro dado muito importante é que as concentrações de ribose no músculo são encontradas em pequenas quantidades. Sendo assim, evidências indicam que as vias para síntese de ribose-5-fosfato e de seus compostos derivados, como o PRPP, apresentam algumas limitações.
Baseado nisso, há a sugestão de que uma maior disponibilidade de ribose poderia resultar em um aumento da formação de PRPP e da taxa de síntese de adenina nucleotídeos. Então, a suplementação de ribose exógena pode ser uma possibilidade no caminho na geração de uma resposta metabólica apropriada para a “ressíntese” de adenina nucleotídeos, pois esse composto possui uma passagem direta para a via da pentose fosfato, sofrendo uma conversão direta para ribose-5-fosfato pela enzima riboquinase, não necessitando da ação da glicose-6-fosfato desidrogenase e 6-fosfogluconato desidrogenase na formação do PRPP. Como resultado, poderia ocorrer aumento na síntese e recuperação de adenina nucleotídeos por uma via mais direta .
Baseado nisso, há a sugestão de que uma maior disponibilidade de ribose poderia resultar em um aumento da formação de PRPP e da taxa de síntese de adenina nucleotídeos. Então, a suplementação de ribose exógena pode ser uma possibilidade no caminho na geração de uma resposta metabólica apropriada para a “ressíntese” de adenina nucleotídeos, pois esse composto possui uma passagem direta para a via da pentose fosfato, sofrendo uma conversão direta para ribose-5-fosfato pela enzima riboquinase, não necessitando da ação da glicose-6-fosfato desidrogenase e 6-fosfogluconato desidrogenase na formação do PRPP. Como resultado, poderia ocorrer aumento na síntese e recuperação de adenina nucleotídeos por uma via mais direta .
NADPH e Glutianona
A importância da via das pentoses fosfato também se relaciona com a redução da glutianona, um redutor imediato de diversos processos celulares e importante eliminador de peróxido de hidrogênio das células. Ao exercer sua ação redutora, os grupos SH de duas moléculas de glutianona são oxidados, formando uma ligação S-S da glutianona dissulfeto, forma não redutora.
A restauração da forma SH da glutianona é obtida por reação com NADPH, tornando-a redutora novamente.
G-S-S-G + NADPH + H ------à 2G –SH + NADP+
A restauração da forma SH da glutianona é obtida por reação com NADPH, tornando-a redutora novamente.
G-S-S-G + NADPH + H ------à 2G –SH + NADP+
Via das Pentoses Fosfato

A partir da observação de que inibidores da glicólise como iodoacetato e fluoreto, em alguns tecidos, não tinham efeito direto na oxidação da glicose, foi sugerida a existência de uma via alternativa para a oxidação da glicose, a vida das pentoses fosfato.
A via das pentoses fosfato é uma via multifuncional, que ocorre principalmente, em tecidos animais, no fígado, na glândula mamária, no córtex da supra renal e no tecido adiposo, onde também ocorre a síntese de ácidos graxos. Bactérias q não possuem aldolase, uma enzima de conversão da frutose 1,6 bisfosfato a diidroxiacetona fosfato e gliceraldeído 3 fosfato usam a via das pentoses para oxidação da glicose. A xilulose 5 fosfato é convertida a gliceraldeído 3 fosfato, havendo a oxidação à piruvato através da 2ª parte da glicólise.
As biossínteses redutoras requerem um poder redutor, encontrado na forma de NADPH. Em tecidos menos ativos em sínteses redutoras, como a produção de ácidos graxos, por exemplo, a via das pentoses é quase ausente.
A via das Pentoses Fosfato produz NADPH e Ribose 5- Fosfato, através de uma etapa oxidativa e uma não oxidativa.
Funções da via das pentoses
a) Permite a oxidação total da glicose em uma série de reações independentes da via glicolítica;
b) É uma fonte de pentoses para a síntese dos ácidos nucleicos;
c) Produz, em uma etapa oxidativa, o NADPH extramitocondrial necessário para a síntese dos lipídios.
d) Seus produtos podem ser isomerizados a intermediários da glicólise
A via das pentoses fosfato é uma via multifuncional, que ocorre principalmente, em tecidos animais, no fígado, na glândula mamária, no córtex da supra renal e no tecido adiposo, onde também ocorre a síntese de ácidos graxos. Bactérias q não possuem aldolase, uma enzima de conversão da frutose 1,6 bisfosfato a diidroxiacetona fosfato e gliceraldeído 3 fosfato usam a via das pentoses para oxidação da glicose. A xilulose 5 fosfato é convertida a gliceraldeído 3 fosfato, havendo a oxidação à piruvato através da 2ª parte da glicólise.
As biossínteses redutoras requerem um poder redutor, encontrado na forma de NADPH. Em tecidos menos ativos em sínteses redutoras, como a produção de ácidos graxos, por exemplo, a via das pentoses é quase ausente.
A via das Pentoses Fosfato produz NADPH e Ribose 5- Fosfato, através de uma etapa oxidativa e uma não oxidativa.
Funções da via das pentoses
a) Permite a oxidação total da glicose em uma série de reações independentes da via glicolítica;
b) É uma fonte de pentoses para a síntese dos ácidos nucleicos;
c) Produz, em uma etapa oxidativa, o NADPH extramitocondrial necessário para a síntese dos lipídios.
d) Seus produtos podem ser isomerizados a intermediários da glicólise
A etapa oxidativa ocorre no sentido da conversão de NADP+ a NADPH, numa reação irreversível catalisada pela lactonase.
A etapa seguinte, não oxidativa, compreende a conversão de ribulose 5 fosfato a ribose 5 fosfato ou xilulose 5 fosfato, pela ação de uma isomerase ou de uma epimerase. Etas pentoses sofrem uma série de conversões, originando intermediários com carbonos variando de 3 a 7 unidade. Essas conversões são catalisadas pelas enzimas transcetolase, que transfere grupos de 2 C e transaldolase, que tranfere grupos de 3 C.
A via das pentoses fosfato ocorre no citossol, assim como a glicólise. Ambas as vias, apesar de possuírem funções diferentes, estão relacionadas por possuírem compostos em comum: a glicose 6 fosfato, a frutose 6 fosfato e o gliceraldeído 3 fosfato.
A reversibilidade das reações da etapa não oxidativa e o compartilhamento de compostos, são os responsáveis pela flexibilidade desta via.
Por exemplo, quando a demanda de NADPH aumenta devido à síntese de ácidos graxos, a via continua a produzir NADPH sem haver acúmulo de ribulose 5 fosfato, pois esse composto pode ser convertido à gliceraldeído 3 fosfato e frutose 6 fosfato, sendo oxidado na via glicolítica. O inverso também pode ocorrer: se a demanda de ribose 5 fosfato, para a síntese de nucleotídeos, por exemplo, for maior que a de NADPH, esse composto pode ser obtido através de compostos pertencentes à via glicolítica.
quinta-feira, 1 de novembro de 2007
Regulaçao da Via das Pentoses
A atividade da via das pentoses varia de acordo com tecido, sendo mais intensa em tecidos que ativam ácidos graxos ativamente, como é o caso do fígado e do tecido adiposo. As duas deidrogenases que participam da etapa oxidativa, convertendo NADP+ a NADPH, são inibidas competitivamente por NADPH.
A utilização da glicose-6-fosfato pela via das pentoses ou pela glicólise vai depender das relações ATP/ADP e NADPH/NADP+ existentes nas células.
Quando a relação ATP/ADP é baixa, a glicose é degradada pela via glicolítica, produzindo ATP. Nessa situação, a síntese de ácidos graxos é baixa e não há consumo de NADPH, sendo a relação NADPH/NADP+ alta, inibindo a via das pentoses.
Mas se a relação ATP/ADP é alta, enzimas chaves da via glicolítica ficam inibidas e a síntese de ácidos graxos é favorecida, havendo um maior consumo de NADPH, diminuindo a relação NADPH/NADP+ e, consequentemente, eliminando a inibição das desidrogenases da via das pentoses.
Portanto, quando a carga energética das células é alta, o consumo de glicose-6-fosfato pela via das pentoses é favorecido. Esta também é ativa quando as taxas glicêmicas são altas. Ou seja, quando a glicemia aumenta, os níveis altos de insulina liberados acarretam, no tecido adiposo, em um aumento da permeabilidade à glicose. Essa condição propicia a síntese de ácidos graxos, que também é estimulada pela insulina.
A utilização da glicose-6-fosfato pela via das pentoses ou pela glicólise vai depender das relações ATP/ADP e NADPH/NADP+ existentes nas células.
Quando a relação ATP/ADP é baixa, a glicose é degradada pela via glicolítica, produzindo ATP. Nessa situação, a síntese de ácidos graxos é baixa e não há consumo de NADPH, sendo a relação NADPH/NADP+ alta, inibindo a via das pentoses.
Mas se a relação ATP/ADP é alta, enzimas chaves da via glicolítica ficam inibidas e a síntese de ácidos graxos é favorecida, havendo um maior consumo de NADPH, diminuindo a relação NADPH/NADP+ e, consequentemente, eliminando a inibição das desidrogenases da via das pentoses.
Portanto, quando a carga energética das células é alta, o consumo de glicose-6-fosfato pela via das pentoses é favorecido. Esta também é ativa quando as taxas glicêmicas são altas. Ou seja, quando a glicemia aumenta, os níveis altos de insulina liberados acarretam, no tecido adiposo, em um aumento da permeabilidade à glicose. Essa condição propicia a síntese de ácidos graxos, que também é estimulada pela insulina.
quinta-feira, 13 de setembro de 2007
Ribose-5-Fosfato
Ribose-5-Fosfato é um pentassacarídeo (açúcar composto por cinco carbonos)com uma ligação do carbono 5 com um fosfato. veja a estrutura da molécula abaixo:

A via das Pentoses Fosfato ou o caminho do Fosfogluconato, uma via alternativa de oxidação de glicose 6-fosfato, produz NADPH e Ribose 5- Fosfato, um precurssor da ribose e desoxirribose necessária na síntese de RNA e DNA respectivamente, além de ser precursor essencial na síntese de nucleótidos e varias coenzimas como o ATP, NADH, FADH2 e coenzima A.
Assinar:
Postagens (Atom)